
種子の発芽と休眠に関わる植物生理
種子は生命の源である。「蒔かぬ種は生えぬ」ということわざがあるように、農業生産において、種子は当然ながら必要不可欠の存在であり、種子は古来より人類の生活と発展を支えた最も重要な財産のうちの1つである。植物にとっても、種子は不適な環境をやり過ごし、遺伝情報を親から子へ伝える媒体として働き、さらには動くことができない母体の代わりに遠くまで散布されて子孫の分布を拡げるという、極めて重要な役割を担っている。今回は種子がどのように形成され、どのように発芽するのか見ていくことにしよう。
〈種子の形成〉
種子の形成のための初期の段階は受精であった(第6回講義資料参照)。今回は、被子植物において、受精が完了したあと、種子が成熟していく過程について説明しよう。
受精卵は、分裂を繰り返して胚球を構成する部分と胚柄を構成する部分に分化する。胚球とは、その名の通り球形の細胞群であり、後に子葉、幼芽(本葉の基になる部分)、胚軸(発芽した時の茎に当たる部分)、幼根(発芽した時の根に当たる部分)に分化する。一方、胚柄は、胚への栄養供給の役割を担っているが、後に退化する。
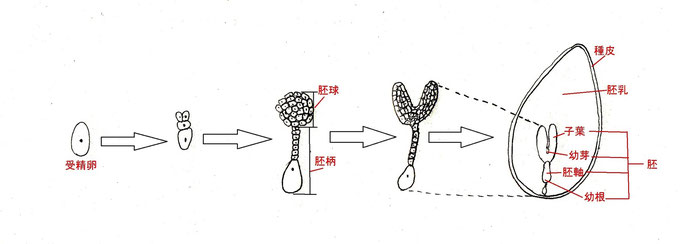
図1 種子形成の過程
胚乳は、発芽の際の養分の供給を担っている組織となる。発芽のための養分が胚乳に蓄えられている種子を有胚乳種子という。一方、発芽のための養分が子葉に蓄えられ、胚乳が退化している種子も存在する。このような種子は無胚乳種子と呼ばれる。前者はイネ科作物、後者はマメ科作物、アブラナ科作物で多く見られる。
種子の成熟や発芽には様々な栄養分が必要となり、種子はこれらの栄養分を親植物から得て、貯蔵している。子葉や胚乳の表皮には、細胞壁の一部が肥厚して大きな内部表面積を持ち、栄養分の効率よい輸送に特化した輸送細胞と呼ばれる細胞が存在する。輸送細胞によって、スクロースなどの低分子の糖類や多糖類である炭水化物、タンパク質、脂肪、リン脂質などが貯蔵される。タンパク質やリン脂質の貯蔵形態であるフィチンはタンパク粒、脂肪はオレオソームという小胞にそれぞれ蓄えられる。また、デンプンはプラスチド(色素体)に貯蔵されるが、セルロースなどデンプン以外の多糖類は細胞壁に蓄えられる。このような貯蔵物質は、発芽の際に分解されて栄養分として用いられる。
種子の各組織の発達が終了すると、細胞分裂が止まり、貯蔵物質が蓄積される。さらにその後、胚が乾燥耐性を獲得して、種子は90%の水分を失い、静止状態になる。この状態の種子が完全に成熟した種子であり、静止種子と呼ばれる。胚の乾燥耐性はアブシジン酸(ABA)という植物ホルモンによって誘導される。
〈休眠とその打破〉
一般に、種子の発芽に必要な条件は水、酸素、そして適切な温度の存在である。小学生時代、発芽のためにはどのような条件が必要か調べるために、暗所や冷蔵庫の中など様々な環境下で種子が発芽するかどうか実験した記憶のある方も多いだろう。静止種子はこれらの3条件を与えるとすぐに発芽する。しかし、多くの場合、植物の種子はこれらの3条件の存在化でも発芽しない。この現象を種子休眠といい、休眠を示す種子を、静止種子に対して休眠種子という。したがって、種子の発芽のためには小学校で習った発芽の3条件に加えて、実はもう1つ、「休眠していない」という条件が必要なのである。種子休眠には植物から採種したときに休眠状態にある一次休眠と、植物から採取した時は非休眠状態にあり、その後の発芽に不適な環境により休眠する二次休眠の2つのタイプに分けられる。また、種子休眠は休眠の原因によって区別されることもある。種皮や胚乳、果皮などが原因になる場合は種皮性休眠と呼ばれる。種皮性休眠では、厚い種皮による酸素供給の抑制や種皮による阻害物質の産生、種皮が固く幼根が突き破れないことなどが原因となる場合が多く、種皮を除去したり、種子表面に傷をつけたりすることによって発芽を誘導することができる。一方、胚自身に休眠の原因がある場合は胚休眠と呼ばれ、これは後述するように植物ホルモンのバランスによって支配されている。
休眠は様々な環境条件によって解除され、これを休眠の打破という。多くの種子で、乾燥によって種子の含水量が低下すると休眠が解除され、この現象は後熟と呼ばれる。一定期間の低温(0~10℃)にさらされなければ休眠が解除されない植物種も存在し、秋に種子を実らせて春に発芽する植物種に多く見られる。また、レタス、ニンジン、ミツバなどのように光によって休眠が打破される植物種も多く、これらの植物の種子は光発芽種子と呼ばれる。(一方で、光によって発芽が阻害される種子は暗発芽種子と呼ばれ、ダイコン、ネギ、カボチャなどの種子が該当する。)
では、胚休眠において、休眠している種子と休眠していない種子では根本的に何が異なっているのだろうか。種子の胚休眠は、植物ホルモンであるABAとジベレリン(GA)の比によって支配されていることが明らかになってきた。ABAの割合が高くなると、休眠が誘導(発芽が抑制)され、GAの割合が高くなると、休眠が解除(発芽が促進)される。ABAは、GAによって誘導されて発芽のための物質生産に使われる酵素の産生を阻害することが分かっている。低温によって休眠が打破される種子では、低温下でABAが分解され、GAが合成される。また、光発芽種子では、光によってGA合成経路の途中で働く酵素の合成が誘導され、最終的にGAの量の増加に結び付く。(光発芽の詳しい仕組みについては文章の最後に参考としてのせているので、興味のある方は読んでほしい。)
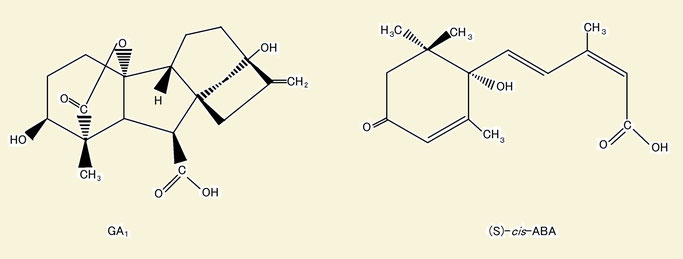
図2:植物体内での活性型GAの1つであるGA1と、活性型ABAである(S)-cis-ABAの構造式
〈発芽の過程〉
発芽の3条件がそろい、かつ休眠が解除された種子はいよいよ発芽の段階に入る。種子の発芽の最初の過程は吸水である。種子は受動的に水を吸い、その重量を増していく。この時期は吸水期と呼ばれている。種子の中の酵素は水によって活性化される。次に、種子は芽や根を形成する細胞を分裂させるための物質やエネルギーを合成する発芽準備期に至る。種子はデンプンやタンパク質、脂肪といった物質を発芽のために蓄えている。このような物質は種子に備わっている消化酵素によって分解され、エネルギー源や細胞構成のための物質になる。デンプンはアミラーゼ及びマルターゼによってグルコース(ブドウ糖)に分解され、呼吸によるATP産生のための基質となる。タンパク質はタンパク質分解酵素によって各種のアミノ酸まで分解され、新たなタンパク質の合成のための原料として用いられる。脂肪はリパーゼによって脂肪酸とグリセリンに分解され、エネルギー源や細胞の構成要素として用いられる。種子の発芽における最終段階は成長期と呼ばれる。成長期では、細胞伸長を促進するオーキシンや細胞分裂を促進するサイトカイニンといった植物ホルモンが合成される。これにより、細胞の伸長・増加が起き、種皮が破られて芽が出る。
休眠の打破にはGAが深くかかわっていると先述した。「休眠していない」ことも発芽のための条件であったため、発芽にはGAが何らかの形で関わっているのではないかと予想される。この予想に対する答えは、イネ科植物の種子で主に明らかになってきた。胚で合成されたGAは、胚乳に放出されて拡散し、種皮と胚乳の間に存在する糊粉層に達する。糊粉層でジベレリンは、α-アミラーゼ遺伝子の転写促進因子であるGA-MYBタンパク質の合成を促進し、結果としてα-アミラーゼの合成が促進される。α-アミラーゼは胚乳に分泌されてデンプンを分解し、得られたグルコースは発芽の際の栄養源として用いられる。
〈発芽率と種子の寿命〉
最後に、発芽率と種子の寿命に関する話をしよう。市販の種の袋には発芽率何%以上という記載がある。発芽率とは、播種した種子のうち何%の種子が正常に発芽するかを示したものである。種を播く際には、発芽率のことを考えて播種量を決めなければならない。また、種袋には種子の有効期限に関する情報も記載されている。種子は発芽能力を永遠に保持できるわけではなく、年月の経過とともに種子に貯蔵された養分は消耗され、発芽率は低下していく。種子の保存には低温・乾燥した環境が適しており、高温多湿の環境は種子の寿命を縮める要因となる。種子の寿命とはすなわち、種子が発芽能力を維持できる期間のことであり、作物種によって大きく異なる。種子の寿命が5年程度と比較的長い種子を長命種子といい、トマトやナス、スイカなどが当てはまる。寿命が3~4年程度の種子は常命種子といい、ダイコンやカブ、ツケナ類、キュウリ、カボチャなどが該当する。寿命が2年程度の作物はレタス、エンドウ、インゲンマメ、ソラマメ、ゴボウ、ホウレンソウなどである。寿命が1年程度しか持たない種子を短命種子といい、ニンジン、タマネギ、ネギ、ラッカセイなどが該当する。
これまでの4回で、採種にかかわる植物生理の基礎知識について説明してきた。次回からはこれまで学んだ知識を用いて、実際の採種の方法を作物別にみていくことにしよう。
参考:光発芽の仕組み
光を感知して発芽するというシステムは、貯蔵養分を少ししか持たない小型の種子にとって、発芽後すぐに光合成を開始できるという点において好都合である。しかし、「光を感知する」という現象と「発芽する」という現象はあまりにかけ離れたものであるように感じられる。植物はどのようにこれらの現象を結び付けているのであろうか。
光発芽種子は、どの波長の光が当たっても発芽するわけではなく、波長が650~680nmの赤色光が照射された場合にのみ発芽が誘導される。植物には光を感知し、形態形成に関与する光受容体と呼ばれる色素タンパク質が数種類備わっている。光発芽にはそのうちのフィトクロムという光受容体によって制御されている。フィトクロムは赤色光照射によってtrans型のPfr、遠赤色光(波長:710~740nm)照射によってcis型のPr型に変換し、この反応は可逆性を持つ。生理活性を持つのはPfr型のフィトクロムであり、すなわち、Pfr型のフィトクロムの割合が多くなっている条件で光発芽種子は発芽する。
Pr型のフィトクロムは細胞質に留まるが、Pfr型に変換されたフィトクロムは核移行性を持つ。Pfr型フィトクロムは、PIF(Phytochrome interacting factor)と呼ばれる転写因子と結合する。PIFはフィトクロムとの相互作用を通じて基本転写因子の活性を制御していると考えられている。つまり、Pfr型フィトクロムは転写因子を介することによって転写の制御を行っている。Pfr型フィトクロムを介して転写誘導を受ける遺伝子の1つにGA3oxがある。この遺伝子は、GA3酸化酵素をコードしており、この酵素は活性型GAであるGA1の前駆体であるGA20の3位の炭素原子に水酸基を導入し、活性型GAを合成する。
このようにして、赤色光の照射という現象が、活性型GAの合成という現象に結び付くのである。GAがどのように種子の発芽を導くかについては本文中で述べたとおりである。本文中ではイネ科植物の種子での例を挙げているが、光発芽種子の代表格であるレタスの種子においても、ジベレリンを与えると、赤色光が照射されなくても発芽が誘導されることが証明されている。
参考文献:
・自家採種入門/中川原敏雄・石綿薫(著)/2009年/農文協
・植物生理学/三村徹郎・鶴見誠二(編著)/2011年/化学同人
・植物生理学 第3版/L.テイツ・E.ザイガー(著)西谷和彦・島崎研一郎(訳)/2004年/培風館
・スクエア最新図説生物/吉田勝利(監修)/2012年/第一学習社
・タネの不思議/田中修(著)/2012年/ソフトバンク クリエイティブ(サイエンス・アイ新書)
・タネの発芽不良の原因と対策/タキイ種苗
http://www.takii.co.jp/tsk/y_garden/autumnsummer/point01/