
受精に関わる植物生理
受精は、植物が種子を作るための最初の段階であり、正常に受精が起きないと採種の成功は望めない。また、さまざまな植物種の受粉の特徴を知ることが、作物の交雑についての理解の助けとなり、品種を正確に保存していくために不可欠である。今回は、被子植物の受精の仕組みと、作物の交雑のしやすさとそれを支配する要因について説明する。
〈被子植物の受精様式〉
まず初めに被子植物の受精について概説しよう。作物のほとんどは被子植物であり、被子植物の受精様式を理解することは、すなわち作物の受精について理解することを意味する。下の図1を見ながら読んでいってほしい。一般に被子植物の花はがく片、花弁、雄ずい(おしべ)、雌ずい(めしべ)の4つの部分からなる。このうち、生殖に直接かかわるのは雄ずいと雌ずいである。雄ずいで作られる花粉の中の精細胞と、雌ずいで作られる胚のうの中の卵細胞が受精して受精卵となる。
雄ずいは葯とそれを支える花糸からなり、葯の中で花粉が産生されている。葯の中にはタペータム組織と呼ばれる花粉母細胞を内蔵する組織があり、このなかにある花粉母細胞(2n)が減数分裂することによって花粉四分子と呼ばれる4つの小胞子(n)が出来る。その後、小胞子は体積が不均等な2分裂を一回行う。大きいほうの娘細胞は栄養細胞、小さいほうの娘細胞は生殖細胞という。生殖細胞は栄養細胞に取り込まれ、細胞壁を消失する(細胞膜は残る)。これが葯から放出される花粉の状態である。
一方、雌ずいは胚珠、子房、柱頭、花柱から構成されており、胚珠が子房に包まれている。胚珠の中には胚のう母細胞(2n)が存在し、これが減数分裂することによって4つの娘細胞(2n)が出来る。これらの娘細胞のうち、3つは退化して1つの胚のう細胞が残る。胚のう細胞は3回の核分裂を起こして8つの核を持った細胞になる。その後、細胞質分裂によって胚のう細胞は3つの反足細胞、1つの中央細胞、1つの卵細胞、2つの助細胞に分裂する(中央細胞は2つの核、それ以外の細胞は1つの核を持つ)。これらをまとめて胚のうと呼ぶ。
受精が起こるには、まず花粉が雌ずいの柱頭に付着する必要がある。柱頭に付着した花粉は、養分や水分を吸収して花粉管を出す。これを花粉の発芽という。花粉管は胚のうを目指して花柱内を伸長していく。多くの植物種で、花粉管伸長のときに花粉内の生殖細胞が分裂し、2つの精細胞を生じる。胚珠に達した花粉管は、胚のうに入って2つの精細胞を放出する。そして、胚のう内の卵細胞と1つの精細胞が受精し(この受精を生殖受精という)、受精卵(2n)が形成される。しかし、これではまだ1つの精細胞が余っている。この余った精細胞は、中央細胞(n+n)と受精し(この受精を栄養受精という)、胚乳(3n)を形成する。つまり、被子植物では2つの受精が起きており、これは被子植物に特有のもので重複受精と呼ばれている。
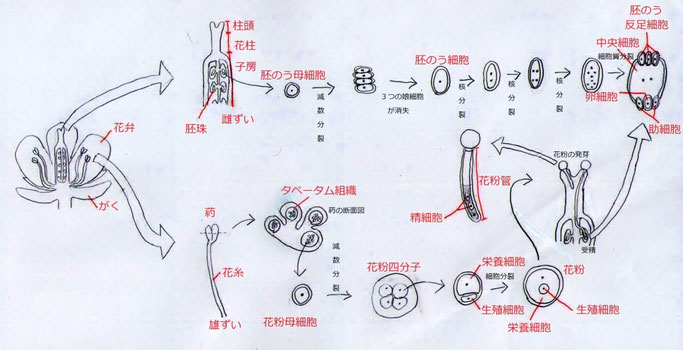
図1 被子植物の花粉・胚のうの形成と受精
ここで、果実の各組織の遺伝子型がどの染色体上の遺伝子に由来しているのか考えてみよう。まず、胚(受精卵が発達したもの)(2n)の遺伝子型は花粉(精細胞)の染色体(n)上の遺伝子と卵細胞由来の染色体(n) 上の遺伝子に由来している。種皮(胚珠を覆う珠皮が成熟したもの)や果実(子房が成熟したもの)は母親由来の組織であり、これらの組織の遺伝子型は完全に母親の染色体(2n)上の遺伝子に支配されており、受精とは無関係である。また、胚乳(3n)の遺伝子型は中央細胞の染色体(n+n) 上の遺伝子と花粉(精細胞)の染色体(n) 上の遺伝子に由来している。したがって、胚乳の遺伝子型は、果実や種皮とは異なり、父親の遺伝子の影響を受ける。この現象をキセニアといい、穀類など子実を利用する作物ではキセニアの影響を大きく受けることがある。例えば、スイートコーンの雌ずいに、ポップコーンの花粉が付着・受精すると、スイートコーンの実にポップコーンの形質が混ざり、品質が低下してしまう。
〈自殖か他殖か 作物の生き残りをかけた適応戦略〉
ここからは作物の受粉相手とその選択による作物の適応戦略(繁殖における有利性を獲得し、種を存続させていくための戦略)について述べていこう。植物の雌ずいの柱頭に付く花粉は様々な個体に由来する。このうち、もっとも近縁であるのは同一個体に由来する花粉であろう。その次に近縁なのは同一品種の他個体に由来する花粉であり、他品種の花粉、異なる種の植物の花粉へと次第に遠縁になっていく。植物はこれらすべての花粉を受精のために受け入れることが出来るわけではない。まず、あまりにも遠縁な種の花粉は受け入れることが出来ない。生物は遠縁な種との受精を避けることによって種の混ざりあいを防いでいる(生殖的隔離)のである。また、もっとも近縁な同一個体の種の花粉については、受け入れることが出来る植物種と、受け入れることが出来ないかあるいは受け入れ可能でもなるべく他個体の花粉を受容するよう工夫している植物種の2つに大別される。前者を自殖性植物といい、同一個体の花粉で受精する自家受精を行って自殖種子を結実させる。後者を他殖性植物といい、他の個体由来の花粉で受精する他家受精を行って他殖種子を結実させる。
まず、自殖性植物についてみていこう。自殖性植物は、それぞれに固有の自家受精を成功させる仕組みを持っている。例えば、アサガオやインゲンでは開花する前にすでに受粉が成立している。イネやコムギでは、開花期間が短く、他家受粉の機会を減らしている。また、トマトのように他家受粉を困難にする花の構造をもつものもある。自殖性植物でも、完全に自殖を行うことは困難で、強い風による花粉の飛散や訪花昆虫による送粉によって他家受粉の割合が高まることがある。植物には近縁な他の個体の花粉を拒む仕組みは備わっていないためである。
一方、他殖性植物は自殖性植物とは逆に、自家受精を避ける仕組みを有している。キャベツ、カブ、ダイコンなどでは両性花(雄ずいと雌ずいの両方をもつ花)において自己の花粉が柱頭に付着してもその後の花粉の発芽や花粉管の伸長が妨げられ、受精が成立しない。この仕組みを自家不和合性という(逆に、自己の花粉を受容する仕組みを自家和合性という)。(自家不和合性の生理学的機構については、記事の最後に参考として載せているので、興味のある人は読んでほしい。) ニンジンやトウモロコシでは、自家和合性を持つが、両性花の中で雄ずいの熟期、すなわち花粉を放出できる期間と雌ずいの熟期、すなわち花粉を受容できる期間がずれている。この仕組みは、雌雄異熟と呼ばれる。ウリ類やトウモロコシでは、自家和合性を持つが、1つの株の中で雄花と雌花が分かれており(雄ずいを持たず雌ずいを持つ雌花、あるいは雌ずいを持たず雄ずいを持つ雄花を単性花という)、自家受粉の確率を低下させている。この仕組みは雌雄異花と呼ばれる。また、ホウレンソウでは、雄花しか持たない雄株と、雌花しか持たない雌株が存在し、両者の間で受粉が起こる。この仕組みを雌雄異株という。(したがって、ホウレンソウを採種する際は多くの株を残さねばならない。少しの株しか残さずに、開花してみて全て雌株あるいは雄株だけだったという事態に陥っても、もう手遅れである。葉野菜として収穫する時期では、外見からまだ雌雄は判別できない。)
雌雄異株では100%の他家受粉が行われているが、雌雄異熟や雌雄異花では他家受粉になるか、自家受粉になるかは確率依存的な問題である。後で述べるように、自家不和合性も条件によっては完全には機能しないことがある。また、先述したように、自殖性作物でも環境条件によっては他家受精を行うことから、自殖性植物と他殖性植物の線引きは明確なものではないことがわかるだろう。自然状態で植物がどの程度他家受精を行うか示した割合を自然交雑率という。自殖性を示す作物を自殖性作物、他殖性を示す作物を他殖性作物と呼ぶが、これらは、実用上は自然交雑率の値で区分されている。一般的に自然交雑率4~5%以下の作物が自殖性作物に区分され、イネ、オオムギ、コムギ、トマト、レタス、ダイズ、インゲン、ラッカセイ、シソなどが挙げられる。また、自然交雑率50%以上の作物は他殖性作物に区分され、ホウレンソウ、トウモロコシ、ダイコン、カブ、ツケナ類、キャベツ、ブロッコリー、キュウリ、カボチャ、メロン、スイカ、ライムギなどが挙げられる。自然交雑率が4~5%より高く、50%より低い作物は部分他殖性作物と呼ばれ、自殖性植物と他殖性植物が混在している。部分他殖性作物としてナス、ワタ、ピーマン、ソラマメ、オクラ、ゴボウ、ニンジン、ネギ、タマネギ、カラシナなどが挙げられる。
では、自殖性作物と他殖性作物はどのような適応戦略を取っているのであろうか。自殖性作物は自らの花粉で受精を行うことによって、個体数が減少しても確実に種を存続できるという戦略をとっている。自殖性作物では親と似通った遺伝情報が継代されていくため、その土地の環境での生育に特化しやすい。したがって、環境の変化に弱いといった弱点も併せ持つ。一方、他殖性作物では、毎世代ごとに他個体と遺伝情報を交換し、遺伝情報の再構成を行っている。このように形質の変わりやすさを特徴として持つことによって様々な環境への適応能力を高めている。しかし、個体数が少なくなると、やがて遺伝子的に近縁な個体間で交雑せざるを得なくなり、この結果、種子の活力が低下し、草勢が弱くなって小型化していく現象がみられる。(次回詳しく述べるが、この現象を近交弱勢という。他殖性作物の中でも、ウリ類のように近交弱勢がほとんど見られないものも存在する。) 作物の採種や品種保存を考える上でもこれらの特徴は極めて重要である。採種に労力がかからないのは自殖性作物であろう。自殖性作物では、異品種との交雑が少なく、他の品種と数メートル隔離しておくだけで十分に品種の維持が可能である。母本(採種に用いる株)も少なくて済む。一方、他殖性作物では交雑を防ぐために多大な労力を費やすこととなる。異品種との交雑は容易に起こりうるし、アブラナ科作物など互いに近縁な作物では異なった作物間(例えばカブとハクサイ)でも雑種が成立する。したがって、訪花昆虫の飛来を防ぐためのネットで囲む、距離的に遠ざける、開花時期をずらすといった物理的・時間的な隔離措置が必要となる。また、品種の維持の面でも他殖性作物では手間がかかる。前述したとおり、他殖性作物は変わりやすさが特徴であり、環境条件によって作物の形質は容易に変化していってしまう。したがって、採種に用いる母本を選抜する際には、元の品種特性を表わしている個体を選ぶという意識を常に持っておかなくてはならない。加えて、他殖性作物には近交弱勢を示すものが多い。このような作物では、母本を多く確保し、様々な個体の花粉が混じり合うようにしておかねばならない。
今回見てきたように、自殖と他殖のシステムは、遺伝的多様性の程度に密接に関連している。そして、遺伝的多様性は植物の形質の強さ、生命力の強さに深く結び付いている。例えば文中でも少し触れたが、行き過ぎた近親交配(同じような遺伝子型の個体間での交配)は、子世代に形質の劣化である近交弱勢を招くことが多い。一方、遠縁の個体同士の交配では、子世代に両親のいずれよりも優れた形質が出る雑種強勢が起こることがある。これは第2回の記事で述べたように、F1品種の作成で広く利用されている性質である。次回は近交弱勢と雑種強勢について詳しく説明していこう。
参考:自家不和合性の仕組み
他殖を実現する仕組みとして、雌雄異花、異花柱性、雌雄異株などが物理的に自家受精を避ける方法を取っているのに対して、自家不和合性では自己の花粉が柱頭につくことは妨げないが、その後の花粉の発芽や花粉管の伸長を妨げるという生理的に自家受精を避ける方法が取られている。自家不和合性の仕組みは、古くから興味が持たれていたが、近年の研究により、植物がどのように自己と非自己を分別しているのか明らかになってきた。
まず、遺伝学的な解析により、自家不和合性の原因遺伝子としてS遺伝子座が見出された。S遺伝子座では、花粉の原因遺伝子と雌ずいの原因遺伝子が隣接して存在する。この2つの遺伝子のセットは近接しているためにあたかも1つの遺伝子のように後代に伝わるため、ハプロタイプと呼ばれる。これらの原因遺伝子は、多くの対立遺伝子を持ち、そのため多様なハプロタイプが存在する。
これまでに、主要な2つのタイプの自家不和合性の仕組みが、主に解析されてきた(図2)。1つは胞子体不和合性と呼ばれるもので、花粉の原因遺伝子が花粉を作った親のDNA(2n)に由来し、アブラナ科などでみられる。もう1つは配偶体不和合性と呼ばれるもので、花粉の原因遺伝子が花粉自身のDNA(n)に由来し、ナス科やバラ科などでみられる。被子植物においては、胞子体は核相2nの植物体を指し、配偶体は核相nの花粉と胚のうを指す。アブラナ科の胞子体不和合性では、葯のタペータム組織(2n)で産生され、花粉表面に結合して情報伝達物質(リガンド)として働いているSCRタンパク質(花粉の原因遺伝子にコードされている)と、柱頭細胞の表面の細胞膜上に存在し、受容体(レセプター)として働いているセリン/トレオニンキナーゼ(SRK)(雌ずいの原因遺伝子にコードされている)が同一のハプロタイプに由来するときに受精が阻止される。(キナーゼとは、リン酸化酵素の意味である。SRKは、自身をリン酸化する自己リン酸化酵素としての作用を持つ。) つまり、同一のハプロタイプに由来するSCRⅠタンパク質とSRKⅠを仮定すると、SCRⅠタンパク質とは異なるハプロタイプに由来するSCRⅡタンパク質を持つ花粉はSRKⅠに結合できず、柱頭細胞表面に付着して発芽を正常に行うが、SCRⅠタンパク質を持つ花粉(すなわち、同一個体由来)は、SRKⅠに特異的に結合し、この結合がシグナルとなって自己リン酸化が起き、最終的に花粉の発芽が阻害される。
一方、配偶体不和合性では、花粉の原因遺伝子がFボックスタンパク質と呼ばれる酵素タンパク質をコードしており、雌ずいの原因遺伝子はRNase(RNA分解酵素)をコードしている。配偶体不和合性ではFボックスタンパク質とRNaseが同一のハプロタイプに由来する場合に受精が阻止される。Fボックスタンパク質は、ユビキチンをRNaseに結合する酵素の役割を担っており、ユビキチンによってマークされたRNaseはタンパク質分解酵素プロテアソームによって分解される。このように、ユビキチンによって印をつけられたタンパク質がプロテアソームによる分解を受ける機構をユビキチン-プロテアソーム系という。また、RNaseは花柱(2n)から分泌され、花粉管の伸長に必要なRNAを分解することによって受精を阻害する。つまり、Fボックスタンパク質の実体であるSLF-SⅠタンパク質とRNaseであるSⅠ-RNaseを仮定すると、SLF-SⅠタンパク質とは異なるハプロタイプに由来するSLF-SⅡタンパク質を持つ花粉では、ユビキチン-プロテアソーム系が正常に働き、RNaseによってRNAが分解されずに済むが、SLF-SⅠタンパク質を持つ花粉(同一個体由来)では、ユビキチン-プロテアソーム系が正常に働かずに、RNaseによってRNAが分解され、花粉管が成長できなくなる。

図2 主要な2つの自家不和合性の仕組み
自家不和合性のシステムは、いつも完璧に働いているわけではなく、条件によっては不完全になることもある。まず、蕾の状態の、未成熟な花では自家不和合性は働かないことが分かった。したがって、蕾のうちに自らの花粉を用いて人工受粉を行うと、自家不和合性を示す作物においても形質の揃った純系の系統を得ることが出来るようになり、F1品種の親世代の育成において重宝される手法となった。また、二酸化炭素濃度が高い環境下においても自家不和合性の機能が失われることが分かっている。蕾の段階での人工授粉は、非常に手間がかかり、大変な作業であった。しかし、採種株を栽培しているハウス内の二酸化炭素濃度をあげ、ミツバチを放って受粉させるとはるかに容易に受粉が出来る。このように、自家不和合性の不活化は、F1品種の育成に大きな恩恵をもたらした。
参考文献:
・自家採種入門/中川原敏雄・石綿薫(著)/2009年/農文協
・植物生理学/三村徹郎・鶴見誠二(編著)/2011年/化学同人
・植物生理学 第3版/L.テイツ・E.ザイガー(著)西谷和彦・島崎研一郎(訳)/2004年/培風館
・山田式家庭菜園教室 スイートコーン/タキイ種苗
http://www.takii.co.jp/tsk/y_garden/spring/sweetcorn/
・植物の特異な自他識別機構を分子レベルで解明/科学技術振興機構・奈良先端科学技術大学院大学
https://www.jsp.go.jp/seika/2011/vol1_003.html
・アブラナ科植物の雄性側の自家不和合性決定遺伝子の優劣性について/新潟大学
http://dspace.lib.niigata-u.ac.jp/dspace/bitstream/10191/18626/1/64
・「一粒の種からのメッセージ」/野口勲
http://noguchiseed.com/hanashi/hitotsubunotane.html